INTRODUCTION
- The human body contains two kinds of glands: exocrine glands and endocrine glands.
- Exocrine glands secrete their products (enzymes) into ducts that carry the secretions into body cavities, into the lumen of an organ, or to the outer surface of the body. Exocrine glands include sudoriferous (sweat), sebaceous (oil), mucous, and digestive glands.
- Endocrine glands (endo-within) secrete their products (hormones) into the interstitial fluid surrounding the secretory cells rather than into ducts. From the interstitial fluid, hormones diffuse into blood capillaries and blood carries them to target cells throughout the body.
- Because of their dependency on the cardiovascular system to distribute their products, endocrine glands are some of the most vascular tissues of the body.
- The endocrine glands include the pituitary, thyroid, parathyroid, adrenal, and pineal glands.
- In addition to that, several organs and tissues are not exclusively classified as endocrine glands but contain cells that secrete hormones. These include the hypothalamus, thymus, pancreas, ovaries, testes, kidneys, stomach, liver, small intestine, skin, heart, adipose tissue, and placenta. Taken together, all endocrine glands and hormone-secreting cells constitute the endocrine system.
- The study of the structure and function of the endocrine glands, the diagnosis and treatment of disorders of the endocrine system is called endocrinology.
Chemically, hormones can be divided into two broad classes:
Lipid Soluble Hormones
Water Soluble Hormones
This chemical classification is also useful functionally because the two classes exert their effects differently.
Lipid-Soluble Hormones
The lipid-soluble hormones include steroid hormones, thyroid hormones, and nitric oxide.
Steroid hormones are derived from cholesterol. Each steroid hormone is unique due to the presence of different chemical groups attached at various sites on the four rings at the core of its structure. These small differences allow for a large diversity of functions.
Two thyroid hormones (T3 and T4) are synthesized by attaching iodine to the amino acid tyrosine. The presence of two benzene rings within a T3 or T4 molecule makes these molecules very lipid-soluble.
Nitric oxide (NO) (Gas hormone) is both a hormone and a neurotransmitter. Its synthesis is catalyzed by the enzyme nitric oxide synthase.
Water-Soluble Hormones
The water-soluble hormones include amine hormones, peptide and protein hormones, and eicosanoid hormones.
Amine hormones are synthesized by decarboxylating (removing a molecule of CO2) and otherwise modifying certain amino acids. They are called amines because they retain an amino group (—NH3). The catecholamines, epinephrine, norepinephrine, and dopamine are synthesized by modifying the amino acid tyrosine. Histamine is synthesized from the amino acid histidine by mast cells and platelets. Serotonin and melatonin are derived from tryptophan.
Peptide hormones and protein hormones are amino acid polymers. The smaller peptide hormones consist of chains of 3 to 49 amino acids; the larger protein hormones include 50 to 200 amino acids. Examples of peptide hormones are antidiuretic hormone and oxytocin; protein hormones include human growth hormone and insulin. Several of the protein hormones, such as thyroid-stimulating hormone, have attached carbohydrate groups and thus are glycoprotein hormones.
Eicosanoid hormones (eicos – twenty forms; oid – resembling) are derived from arachidonic acid, a 20-carbon fatty acid. The two major types of eicosanoids are prostaglandins and leukotrienes. The eicosanoids are important local hormones, and they may act as circulating hormones as well.
HORMONE ACTIVITY
The Role of Hormone Receptors
Although a given hormone travels throughout the body in the blood, it affects only specific target cells. Hormones, like neurotransmitters, influence their target cells by chemically binding to specific proteins called receptors. Only the target cells for a given hormone have receptors that bind and recognize that hormone. For example, thyroid-stimulating hormone (TSH) binds to receptors on cells of the thyroid gland, but it does not bind to cells of the ovaries because ovarian cells do not have TSH receptors.
Receptors, like other cellular proteins, are constantly being synthesized and broken down. Generally, a target cell has 2000 to 100,000 receptors for a particular hormone. If a hormone is present in excess, the number of target-cell receptors may decrease an effect called down-regulation. Downregulation makes a target cell less sensitive to a hormone. In contrast, when a hormone is deficient, the number of receptors may increase. This phenomenon is known as up-regulation which makes a target cell more sensitive to a hormone.
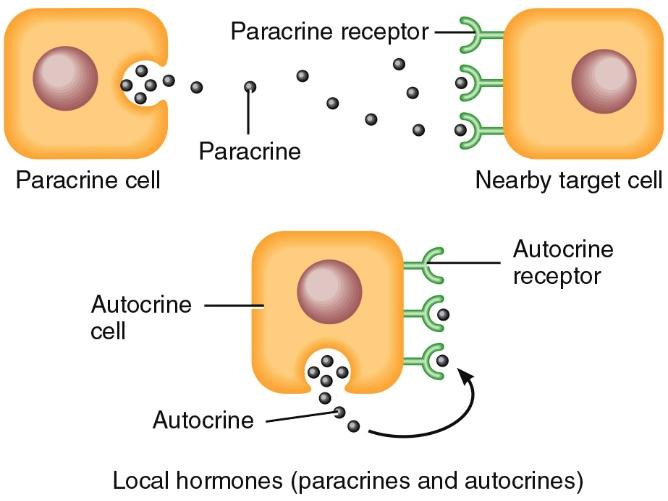
Circulating and Local Hormones
Most endocrine hormones are circulating hormones. They pass from the secretory cells that make them into the interstitial fluid and then into the blood.
Other hormones, termed local hormones, act locally on neighbouring cells or on the same cell that secreted without first entering the bloodstream. Local hormones that act on neighbouring cells are called paracrine (para- beside or near), and those that act on the same cell that secreted them are called autocrine (auto- self).
One example of a local hormone is interleukin-2 (IL-2), which is released by helper T cells (a type of white blood cell) during immune responses. IL-2 helps activate other nearby immune cells, a paracrine effect. But it also acts as an autocrine by stimulating the same cell that released it to proliferate. This action generates more helper T cells that can secrete even more IL-2 and thus strengthen the immune response.
Another example of a local hormone is the gas nitric oxide (NO), which is released by endothelial cells lining of blood vessels. NO cause relaxation of nearby smooth muscle fibres in blood vessels, which in turn causes vasodilation (increase in blood vessel diameter). The effects of such vasodilation range from a lowering of blood pressure to erection of the penis in males.
Local hormones usually are inactivated quickly; circulating hormones may linger in the blood and exert their effects for a few minutes or occasionally for a few hours. In time, circulating hormones are inactivated by the liver and excreted by the kidneys.

Hormone Transport in the Blood
Most water-soluble hormone molecules circulate in the watery blood plasma in a “free” form (not attached to other molecules), but most lipid-soluble hormone molecules are bound to transport proteins that are synthesized by cells in the liver
Transport proteins have three functions:
They make lipid-soluble hormones temporarily water-soluble, thus increasing their solubility in the blood.
They slow the rate of hormone loss in the urine by retard the passage of small hormone molecules through the filtering mechanism in the kidneys.
They provide a ready reserve of hormones, already present in the bloodstream.
MECHANISM OF HORMONE ACTION
The response to a hormone depends on both the hormone and the target cell. Various target cells respond differently to the same hormone. Such as insulin stimulates the synthesis of glycogen in liver cells and the synthesis of triglycerides in adipose cells.
The response to a hormone is not always the synthesis of new molecules, it can change the permeability of the plasma membrane, alter the rate of specific metabolic reactions and can cause contraction or relaxation of muscles.
However, a hormone must first “announces its arrival” to a target cell by binding to its receptors. The receptors for lipid-soluble hormones are located inside target cells and the receptors for water-soluble hormones are part of the plasma membrane of target cells.
Lipid soluble hormones, including steroid hormones and thyroid hormones, bind to receptors within target cells. Their mechanism of action is as follows:
A free lipid-soluble hormone molecule diffuses from the blood, through interstitial fluid, and through the lipid bilayer of the plasma membrane into a cell.
Inside the target cell, the hormone binds to and activates receptors located within the cytosol or nucleus. The activated receptor–hormone complex then alters gene expression: It turns specific genes of the nuclear DNA on or off.
As the DNA is transcribed, new messenger RNA (mRNA) forms, leaves the nucleus and enters the cytosol. There, it directs the synthesis of a new protein, often an enzyme, on the ribosomes.
The new proteins alter the cell’s activity and cause the responses typical of that hormone.

Because amine, peptide, protein, and eicosanoid hormones are not lipid-soluble, they cannot diffuse through the lipid bilayer of the plasma membrane and bind to receptors inside target cells. Instead, water-soluble hormones bind to receptors that protrude from the target-cell surface. The receptors are integral transmembrane proteins in the plasma membrane. When a water-soluble hormone binds to its receptor at the outer surface of the plasma membrane, it acts as the first messenger which causes the production of a second messenger inside the cell. One common second messenger is cyclic AMP (cAMP). Neurotransmitters, neuropeptides, and several sensory transduction mechanisms also act via second messenger systems.
The action of a typical water-soluble hormone occurs as follows:
A water-soluble hormone (the first messenger) diffuses from the blood through interstitial fluid and then binds to its receptor at the outer surface of a target cell’s plasma membrane. The hormone-receptor complex activates a membrane protein called a G protein. The activated G protein in turn activates adenylate cyclase.
Adenylate cyclase converts ATP into cyclic AMP (cAMP) in cytosol because the enzyme’s active site is present on the inner surface of the plasma membrane.
Cyclic AMP also known as the second messenger, activates one or more protein kinases, which may be free in the cytosol or bound to the plasma membrane. A protein kinase is an enzyme that phosphorylates (adds a phosphate group to) other cellular proteins (such as enzymes). The donor of the phosphate group is ATP, which is converted to ADP.
The activated protein kinases phosphorylate one or more cellular proteins. Phosphorylation activates some of these proteins and inactivates others, rather like turning a switch on or off.
Phosphorylated proteins in turn cause reactions that produce physiological responses. Different protein kinases exist within different target cells and within different organelles of the same target cell. Thus, one protein kinase might trigger glycogen synthesis, a second might cause the breakdown of triglyceride, and third may promote protein synthesis, and so on. As noted, phosphorylation by a protein kinase can also inhibit certain proteins. For example, some of the kinases unleashed when epinephrine binds to liver cells inactivate an enzyme needed for glycogen synthesis.
After a brief period, an enzyme called phosphodiesterase inactivates cAMP. Thus, the cell’s response is turned off unless new hormone molecules continue to bind to their receptors in the plasma membrane.

The responsiveness of a target cell to a hormone depends on
(1) The hormone’s concentration in the blood
(2) The abundance of the target cell’s hormone receptors
(3) Influences exerted by other hormones.
PERMISSIVE EFFECT: A target cell responds more vigorously when the level of a hormone rises or when it has more receptors (upregulation). Besides, the actions of some hormones on target cells require simultaneous or recent exposure to a second hormone. In such cases, the second hormone is said to have a permissive effect.
For example, epinephrine alone only weakly stimulates lipolysis (the breakdown of triglycerides), but when small amounts of thyroid hormones (T3 and T4) are present, the same amount of epinephrine stimulates lipolysis much more powerfully.
SYNERGISTIC EFFECT: When the effect of two hormones acting together is greater or more extensive than the effect of each hormone acting alone, the two hormones are said to have a synergistic effect.
For example, the normal development of oocytes in the ovaries requires both follicle-stimulating hormone from the anterior pituitary and estrogens from the ovaries. Neither hormone alone is sufficient.
ANTAGONISTIC EFFECTS: When one hormone opposes the actions of another hormone, the two hormones are said to have antagonistic effects.
An example of an antagonistic pair of hormones is insulin, which promotes the synthesis of glycogen by liver cells, and glucagon, which stimulates the breakdown of glycogen in the liver.
CONTROL OF HORMONE SECRETION
The release of most hormones occurs in short bursts, with little or no secretion between bursts. When stimulated, an endocrine gland will release its hormone in more frequent bursts, increasing the concentration of the hormone in the blood. In the absence of stimulation, the blood level of the hormone decreases. Regulation of secretion normally prevents overproduction or underproduction of any given hormone.
Hormone secretion is regulated by
(1) signals from the nervous system
(2) chemical changes in the blood and
(3) by other hormones.
Good night
ReplyDelete